|

Abstract
Table
of Contents
Part I
Part II
Part III
Part IV
Appendixes
|
Part II: The Seed of Woman
Chapter 18
The Seed Of The Woman And The Seed
Of The Man
I will put enmity between thee (Satan)
and the woman,
between thy seed
and her seed;
it shall bruise thy head.
(Genesis 3:15)
When Darwin formulated
his theory of Natural Selection, it seemed obvious to him, and
he easily persuaded many of his contemporaries, that any living
thing which acquired a particular advantage over its competitors
during its life time, would automatically pass that benefit on
to its young by inheritance. Thus by a process akin to compound
interest, the gains of each new generation were added to those
of the last and linear progress in the development of higher
and higher forms of life was guaranteed.
Any such supposed advantage accruing
to an individual as the result of life experience is generally
referred to as an Acquired Characteristic. For example, the man
who becomes a blacksmith and develops tremendous arm and shoulder
muscles would automatically endow his children with a superior
physique, even though he himself may have been something of a
weakling as a child. One of the strongest advocates of the inheritance
of such acquired characteristics was the French naturalist Jean
Baptiste Lamarck (1744�1829).
The fact seemed obvious and essential
if any progress was to be made in improving the species of animals
of particular value to man,
 pg.1
of 29 pg.1
of 29 
and superficially there
seemed to be a great deal of evidence in support of it. Darwin
was confident that he had the mechanism he needed to make his
hypothesis workable even when applied to human society. But towards
the end of the last century it was becoming increasingly apparent
that Lamarck was mistaken, that acquired characteristics are
either not inherited at all or so rarely that the assumed linear
progress of life through geological ages could not be attributed
to the operation of any such mechanism.
Moreover, it quickly became obvious that
there were many examples in human history of "mutilations"
persistently practiced by men upon their fellows for centuries
which nevertheless were not inherited. Chinese girls, from time
immemorial, had their feet tightly bound because it was felt
that small feet added to a woman's beauty, yet Chinese babies
were still born with normal feet (187)
One of the most famous experiments of
this nature in which an attempt was made to demonstrate whether
mutilation of the parents could lead to defective offspring,
was carried out by August Weismann (1834�1914), Professor
of Biology at the University of Freiburg and Breisgau, whose
chief interest was in Embryology. Weismann cut off the tails
of hundreds of rats, generation after generation, but never succeeded
in getting any baby rats born without tails. Some witty individual
with a literary background and inspired by Shakespeare observed,
"There is a divinity doth shape their ends, rough hew them
though we may!"
Weismann's conclusion, based almost entirely
upon reflection rather than on experiment, was that every body
carried within itself in some kind of concentrated form a deposit
of hereditary substance which he termed "the germ plasm."
(188) This was
the reservoir of specialized material out of which the elements
of the next generation would be derived. He believed that
unless external influences broke through the defenses surrounding
this reservoir of germ plasm, there would be no transmission
of any characters acquired during the life time of the parent.
Such characters affected only the body of the parent and not
his or her germ plasm. If such characters acquired during life
did show up in the offspring, it must be presumed that somehow
the influence of these characters had penetrated the defenses
and reached the germ plasm. He even went so far as to hypothesize
that the germ plasm was particulate in nature, the particles
each in some way being carriers of an inheritable factor. He
knew nothing about genes at the time, a fact which makes his
insight all the more remarkable.
Commenting on this apparent resistance
to change which is built into all living things, Sir Julian Huxley,
in 1938, said, (189)
Can the hereditary constitution be permanently changed by
the environment? It is clear that theoretically it should be
possible to
187. See Notes at the end of this chapter
(page 19).
188. Weismann, August, Essays Upon Heredity and Kindred Biological
Problems, translated by E. B. Poulton, S. Schonland and A.
E. Shipley, Oxford University Press, 1889, vol.1, p.419ff.
189. Huxley, Sir Julian, "Inheritance of Acquired Characteristics"
in Essays in Popular Science, Penguin, 1938, p.36, 37.
pg
2 of 29
induce such changes. The hereditary constitution
is seen to be something material which only our lack of knowledge
prevents us from defining chemically; and as such it must be
possible for us to alter it. The remarkable fact, however, is
its stubbornness in resistance to alteration.
Sixty-nine generations of flies
bred in the dark � and yet no alteration in their eyes or
their instincts with regard to light. Ninety generations in an
attempt to raise their resistance to heat by acclimatization
and selection � without results. Indefinite time spent by
dandelions in the lowlands not preventing their immediately reacting
to mountain conditions by changing size and form and proportion
� and vice versa on replanting from mountain to plain. .
. .
In spite of all the work that has been
done, we have only established the very definite certainty that
to a great many apparently outward influences the germ plasm
is quite unresponsive.
Professor Raymond
Pearl of Johns Hopkins University, after outlining experiments
which involved the controlled breeding of over 300 generations
of one species of fly, concluded: (190)
[This is] perhaps the longest
bit of controlled breeding ever carried out with the result in
each successive generation carefully observed and precisely recorded.
Allowing 30 years as a round figure for the average duration
of a human generation, the time equivalent in human reproduction
of this experiment would be of the order of 9000 years . . .
considerably longer than the total span of man's even dimly recorded
history.
The objective
of this experiment was, of course, to see whether it would be
possible in any way to influence the germ plasm by various manipulations
of the environment. Raymond Pearl summed up the situation by
saying, "the demonstration of the inherent stability of
the genic mechanism of heredity that this experiment has given
is extremely impressive." (190)
Now the explanation for this negative
conclusion is owing to a large extent to the work of Weismann.
Quite early in his professional career as an embryologist, he
began to find that he could no longer continue his research in
developmental physiology due to failing eyesight which seriously
restricted his use of a microscope. As a result, he turned to
the theoretical aspects of his subject � with remarkably
beneficial consequences in terms of our subsequent understanding
of the earliest stages in the development of the fertilized ovum.
His basic conclusions have since been "substantiated to
a surprising degree by the work in genetics in succeeding years,"
as Robert Briggs and Thomas King have observed. (191)
The fertilization of the ovum by a spermatozoon
initiates a process of development in which the seed begins to
multiply until a certain number of cells are formed, all of which
appear to share the constitution
190. Pearl, Raymond, "Biology and Human
Trends," Smithsonian Institute Report for 1935, Smithsonian
Institute, Washington, D.C., Publication #3364, Washington,
1936, p.331.
191. See in Notes at the end of this chapter (page
20)
pg.3
of 29
and totipotency * of
the ovum itself. Then, for reasons which are only just now beginning
to be understood, further division of some of these cells is
accompanied by a change in their constitution which may be due
to their reduced size or their orientation with respect to the
rest of the cells, or to chemical alteration, or to internal
re-organization related to the time lapsed since the process
of cleavage began. (192)
It has been proposed by Christian P. Raven that the cytoplasm
has begun to develop a complicated spatial structure due to a
re-orientation of its contents as it ages or due to some genuinely
new structures arising as a result of chemical reactions taking
place in the egg. At any rate, the various parts of the egg,
which were all alike before, now begin to show differences in
chemical composition. (193) Weismann rightly surmised that some such change signalled
the beginning of the emergence of body cells, the rest
of the original cells meanwhile preserving their character specifically
as germ cells. Thus the germ cells give rise to the germ
cells or seed of the next generation: while the body cells give
rise to the organs of reproduction which will house them and
see to their ultimate fertilization, as well as to the body of
which these organs form a part. The body cells merely serve as
the arena in which this reproductive process is brought to maturity.
Thus is carried forward the germ plasm from generation to generation
in an unbroken chain. The body cells are built out of, and
by, the germ cells or germ plasm; the germ plasm is not built
out of the body. It was this basic hypothesis which was perhaps
Weismann's most important contribution. It is commonly termed
"the continuity of the germ plasm".
The following diagram (Fig. 7)
may help to show what is really involved here, underscoring the
rather humbling fact that the body is
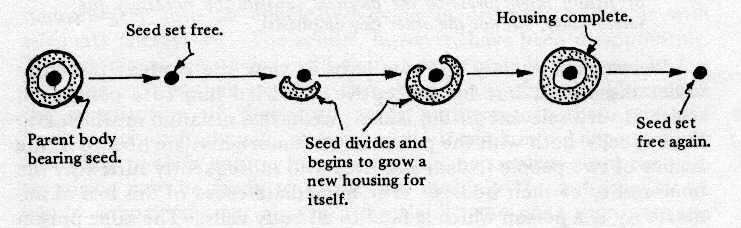
Fig. 7. Seed gives rise to Seed.
* Totipotency: the ability of a cell to multiply
into a whole organism.
192. See Note at the end of this chapter (page 20).
193. Raven, Christian P., An Outline of Developmental Physiology,
translated by L. de Ruiter, New York, McGraw Hill, 1954, p.62.
pg.4
of 29
almost incidental, being
merely the housing for the seed; whereas the seed is the only
truly continuing element.
Generation after
generation, bodies die and return to the dust, but the seed continues
in an unbroken line reaching, in fact, uncorrupted in the woman
from Adam to Mary and of course it still continues to perpetuate
itself.
The simplest way of explaining how physical
death has passed upon all men through man is to assume that in
natural generation the corruption which finally overwhelms the
bodies of men and women alike is introduced to the ovum via the
male seed but does not actually take effect until the stage of
embryonic development has been reached at which these differentiated
body cells have begun to form. The mortogenic factor has apparently
had no influence upon the germ plasm of the woman but only upon
the germ plasm of the man. By contrast, it does have its deadly
influence upon the differentiating body cells of BOTH the male
and female embryo once the body cells begin to form by diverging
in their constitution from the germ plasm. In short, the germ
plasm of the male, and the bodies of both males and females,
are mortalized. But the germ plasm of the female remains immortal.
That such a mechanism might be responsible
for changes in cells subsequently derived from the germ plasm
was suggested by Weismann. In 1881 he wrote: (194)
It may be objected that cells of which
the ancestors possessed the power of living forever could not
become potentially mortal either suddenly or gradually, for such
a change would contradict the supposition which attributes immortality
to their ancestors and to the products of their division. This
argument is valid, but it only applies so long as the
descendants retain their original constitution. As soon as
the two products of fission of a potentially immortal cell acquire
different constitutions by unequal fission, another possibility
arises. It is conceivable that one of the products of fission
might preserve the physical constitution necessary for immortality,
but not the other [my emphasis]
Weismann certainly
did not have in view the context presently under discussion,
but his perceptive mind led him to a conclusion which is very
relevant to the issue. We have a situation in which two lines
of cells, both with the potential of immortality, are housed
in the bodies of two people (Adam and Eve) who subsequently surrender
the immortality of their bodies. The immediate cause of this
loss of immortality is a poison which is fatal to all body
cells. The same poison also proves fatal to the male spermatozoa
which will later be generated out of this germ plasm. Unlike
the oocytes in the female which are already formed at the time
of birth, spermatozoa are
194. Weismann, August, "Upon the Eternal
Duration of Life" in Essays Upon Heredity and Kindred
Biological Problems, translated by E. B. Poulton, S. Schonland
and A. E. Shipley, Oxford University Press, 1889, vol.1, p.139.
pg.5
of 29
manufactured throughout
the adult life of the male. They are apparently susceptible to
the influence of body cells, especially those of the tissues
which generate them.
When any attempt is made artificially
to promote self-replication and further development of a single
spermatozoon, the results are negative. The sperm are not viable
for more than a few days unless fused with the ovum. But when
the female ovum is treated suitably (at least in the animal world
below man), it may develop into a mature organism. It is capable,
therefore, of replicating itself indefinitely, even in the absence
of fusion with a spermatozoon. When this is observed in nature
it is referred to as parthenogenesis, meaning essentially
virgin conception leading to virgin birth. The fact of parthenogenesis
is clearly established for the female seed: (195) the same cannot be said to have been observed for
the male seed.
The ovum is, in fact, a unicellular organism.
And by virtue of its ability to replicate itself indefinitely
under appropriate conditions, it must be assumed to have retained
the same kind of physical immortality which other unicellular
organisms (like amoeba or paramecia, for example) still enjoy.
By contrast, although the individual spermatozoon has all the
appearance of a highly active and extremely complex unicellular
organism, it does not have the power to replicate itself, and
therefore does not enjoy a like physical immortality. It does
not behave like a unicellular organism.
Sometimes it is argued that the proportion
of cytoplasm surrounding the nucleus of the male spermatozoon
is too small to supply it with the energy reservoir necessary
to enable it to replicate itself. As a consequence, the nucleus
cannot survive for more than 24�36 hours or so, unless it
is fused with the ovum: it simply runs out of energy. The female
seed has several thousand times (196) as much food available to supply the energy for cleavage
because it is so much larger, though the nucleus itself
is no larger than in the male seed. To test whether this is true,
the male nucleus has been experimentally transferred by microsurgery
into an enucleated ovum in order to supply it with adequate energy.
(197) The results,
however, have been disappointing. The spermatozoon nucleus still
cannot perpetuate itself beyond a few divisions. It is thus apparent
that there is a profound difference in the constitution of these
two seeds in respect to their potential immortality. Both will,
or course, die if not housed appropriately according to their
nature, but whereas the mammalian ovum can with surprising ease
be made to divide and multiply (198) and grow into a whole animal (though always a female)
(199), the male
spermatozoon cannot.
Whatever the nature of the defect
in the male seed brought about by the entrance of the poison,
it is almost certainly the channel by
195. See Notes at the end of this chapter
(page 21).
196. See Notes at the end of this chapter (page
22)
197. See Notes at the end of this chapter (page
22).
198. See Notes at the end of this chapter (page
23).
199. See Notes at the end of this chapter (page
24).
pg.6
of 29
which the mortogenic
factor is introduced into the ovum at the time of conception,
even though the effect itself is not felt in the presumptive
organism until the multiplying cells begin the process of differentiation
for the express purpose of forming body cells. (200)
Now Weismann set forth this theory
of the "continuity of the germ plasm" in the following
way (Fig. 8):
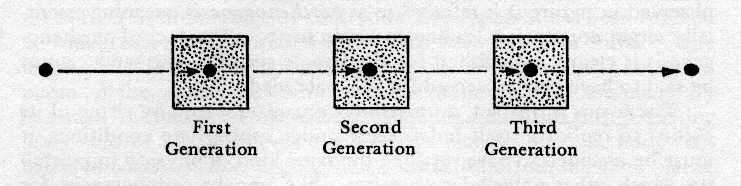
Fig. 8. Each seed gives rise to nother seed and
to the body which houses it.
With remarkably
few modifications, his conclusions have stood the test of time.
(201) His thinking
triggered the somewhat facetious remark which is often made to
freshmen students when they are first introduced to these concepts:
"The hen is merely the egg's way of laying another egg."
The thought is a depressing one if man himself is viewed merely
as a plaything of Nature, a by-product of a process bent upon
a blind course of species improvement without respect to the
worth of the individual. For the individual becomes simply one
stage in an entirely impersonal process. As Kenneth Walker put
it rather effectively: (202)
All that the somatic cells,
which form the main bulk of man's body are really called upon
to do is to provide a refuge in which the immortal cells . .
. can find temporary lodging and sustenance. It is a little bit
discouraging to our self esteem to be looked upon as merely useful
wallets for conveying the valuable germ plasm down the ages.
It is, then,
a simple fact that the body does not generate the ova (in which
case the ova would inevitably have become heir to the defect
of the body) but the ovum generates the body. As Professor A.
S. Pearse put it, "through a series of divisions a germ
cell gives rise to a body or soma and to new germ cells. The
latter, and not the body, give rise to the next generation."
(203) It appears
that this mechanism is by no means limited to human generation.
It is a phenomenon of very wide occurrence in sexually reproductive
organisms below man. Alfred Huettner describes this process as
it occurs in the roundworm: (204)
200. B. Bacetti and B. A. Afzelius, in their
definitive study of the sperm cell [op. cit., ref. #122],
have remarked specifically upon the very high percentage of defective
sperm. See further on this Note #217 at the end of this chapter
(page 27).
201. See Notes at the end of this chapter (page
24).
202. Walker, Kenneth, Meaning and Purpose, London, Penguin,
1950, p.63.
203. Pearse, A. S., General Zoology, New York, Henry Holt,
1930, p.379.
204. Huettner, Alfred F., Fundamentals of Comparative Embryology
of the Vertebrates, New York, Macmillan, revised edition,
1968, p.6,7.
pg.7
of 29
In certain
forms, as for example in Ascaris, the single primordial
germ cell is set aside in the second cleavage of the ovum, and
while this cell continues to divide it does so at a retarded
rate. The other cells keep on cleaving at their usual rate and
eventually form the body or soma of the animal, while the retarded
germ cells become enclosed in the body to develop the gonads.
. . . It is the function of these somatic cells to carry the
germ plasm and nourish and protect it. The somatoplasm has to
die some time and revert to the inorganic world. The germ plasm,
however, is protected by and parasitic on the somatoplasm and
is immortal so long as, during the life of the individual, one
or a number of cells from the germ plasm (the gonad) with the
total number of determinants is liberated and becomes activated
by fertilization or by parthenogenesis and develops again into
germ plasm (gonad) and somatoplasm (body) in the next generation
and so ad infinitum.
Sir Charles
Sherrington has a beautiful free-flowing passage describing this
situation. Coming as it does from a man whose life was spent
largely in medical research and whose reputation was international,
his words so eloquently expressed are doubly worth pondering.
Nor has recent research required their modification since they
were penned. As applied to human beings his statement is still
correct. He wrote: (205)
In its earliest stages the embryo's
cells are not notably different one from another. Later . . .
they become so in spite of being by descent all members of one
family. . . .
To this there seems at first sight
one exception, one cell-type which, out of all the myriads, alone
remains its original self and does not specialize. It retains
the old original nature of the ancestral cell. Its sisters and
their progeny pass on through chains of metamorphoses to form
a world of different shapes and activities. But this one persists
still unmodified and true to its own primitive forebear. . .
.
All its sisters with their flights
into far-fetched specializations, including the brain with its
mysteries of mind, are powerless to produce again a germ such
as they sprang from. From no one of them all, let them be ever
so human, can any fertilization produce their like again in the
shape of man or human child. . . .
From the old ancestral cell one
narrow derivative line of descendants, nested in the rest of
the immense collateral progeny, retains its original germinal
and general nature: and this, even, has to ripen. Significantly
enough, it then sets itself free from all the others. And so
on from generation to generation.
It is therefore clear
that these germ cells constitute a very slender thread in the
continuity of immortality, for the initial cell (the fertilized
ovum) retains its own identity for between two and five doublings
(depending upon the species) before differentiated cells begin
to appear which can no longer be considered as part of the original
germ plasm. These few pure germ cells will continue to replicate
themselves
205. Sherrington, Sir Charles, Man on His
Nature, Cambridge, 1950, p.94, 95.
pg.8
of 29
in isolation, though
at a much slower rate, but for a short interval of time the stream
of immortality is entrusted to a tiny handful of cells.
The expert in these matters will
not need elaboration of this circumstance but the layman may
find it helpful, in visualizing how the ovum provides for its
own continuity, to have the following summary statement.
The sperm penetrates the
ovum and shortly thereafter the ovum begins to divide into two
cells, then each of these divide again and we have four cells.
Shortly, there are eight and then sixteen, and so it grows into
a ball of cells called a morula. A fluid-filled hollow
develops and the whole growing mass assumes the form of a kind
of thin-shelled ball like an orange peel without the orange inside.
This is the blastocyst stage. The blastocyst then collapses on
itself, looking rather like an air-filled ball with a small hole
in it that has been stepped on and stays that way. In time, various
parts of the structure begin to develop differently and the foundations
for the reproductive system begin to emerge in what is called
the genital ridge, which is approximately where the rubber ball
indented and tended to close up the fold. Meanwhile, the germ
cells have kept themselves apart in one place, multiplying slowly.
Once the blood vessels and a kind of circulation system is in
operation, these germ cells which have still retained their integrity
migrate via the vascular system by a form of amoeboid motion
towards the area of the genital ridge. When the gonads finally
form in this area, they are invaded by the germ cells which then
take up residence there. The gonads themselves now begin to develop
as testes or ovaries, depending more or less on the chromosomal
sex of the original germ cells.
It will be remembered that the
presence of the X or Y chromosome determines whether the medulla
or the cortex of the gonads will grow at the expense of the alternative
and therefore whether testes or ovaries will form. These glands,
once the decision is made as to which they shall become, begin
to secrete hormones which act upon the maturing fetus to cause
the appropriate internal reproductive organs to form, and later
the appropriate external genitalia. By full term, the female
fetus has its internal reproductive organs (ovaries, fallopian
tubes, uterus, etc.) all essentially complete and supplied with
a full quota of oocytes which, one by one, will later mature
and be released as fully prepared ova throughout the whole fertile
period of the woman's life.
If one wanted to think in
purely biological terms, one might now convert a previous popular
observation to read, "the woman is merely the ovum's way
of creating another ovum." This is so because the seed of
the woman perpetuates itself in a very special way � as we
shall see in the next chapter.
pg.9
of 29
Now various figures are given for the number of "pure"
germ cells (from 16 to 32) after which further cleavage results
in the appearance of the first somatic cells. (206) Of Ascaris megalocephala, Alfred Kuhn says
that the first somatic cells appear at the fourth cleavage
when there are 16 cells present. The experimental evidence is
sometimes contradictory but it suggests a basic pattern: the
differentiation of mortal cells from immortal cells is made very
early in development. (207)
Thus the germ cells do not
continue their uninterrupted development for very long. They
soon throw off differential cells which signal the development
of body tissues. But at the same time they preserve themselves
as a reservoir of germ plasm throughout the life of the organism.
The subsequent history of
the developing germ plasm in the female is, however, different
from that of the male germ plasm. As J. Money and A. A. Ehrhardt
observe: (208)
At birth the normal pair of
ovaries is said to have between 300,000 and 400,000 ova, of which
approximately 300 to 400 will eventually go through the process
of ovulation. The billions of sperm produced by the testes are
not all present at birth as ova are believed to be, but are produced
new throughout life.
Now this basic
fact of the continuity of the germ plasm has been set forth diagrammatically
in a number of ways by various authorities since Weismann's time.
In a paper entitled, "The Third Stage in Genetics,"
Donald Michie has the following figure showing in a simplified
way two opposing views of the fate of the germ plasm from
generation to generation. (209) I have modified his drawing slightly in order to
make its meaning more self-evident to those who find diagrams
difficult.
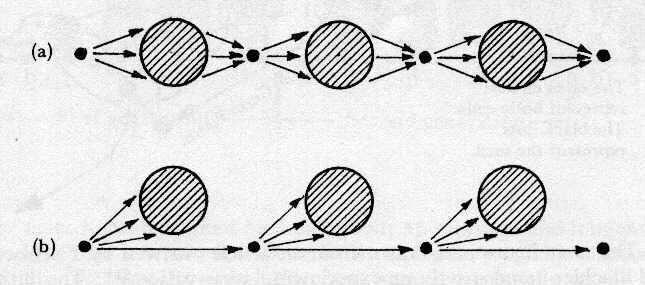
Fig. 9. Two opposing views of the fate of the seed as
it passes from generation to gneration.
206. See Notes at the end of this chapter (page 25).
207. Kuhn, Alfred, Lectures in Developmental Physiology, translated
by Roger Milkman, New York, Springer-Verlag, 1971, p.481.
208. See Notes at the end of this chapter (page
25).
209. Michie, Donald, "The Third Stage in Genetics",
in A Century of Darwin, edited by S. A. Barnett, London,
Heineman, 1958, p.57.
pg.10
of 29
In Fig. 9 (a) the germ seed gives rise to a body which
then gives rise to a germ seed. The latter then gives rise to
a second generation body which in turn generates a second generation
seed. And so the process goes on indefinitely. The important
thing is that in this view the body really is giving rise
to the seed. It is a view which was commonly held until the time
of Weismann. In Fig. 9 (b) the situation is really quite different,
for the initial germ seed gives rise to the next germ seed and
to a body, the germ seed and the body thus generated being
almost independent entities. This is not quite true and
to this extent the diagram is unsatisfactory except in so far
as it tends to point up the two different concepts very nicely.
It is not quite true in so far as the seed is dependent
upon the body to house it.
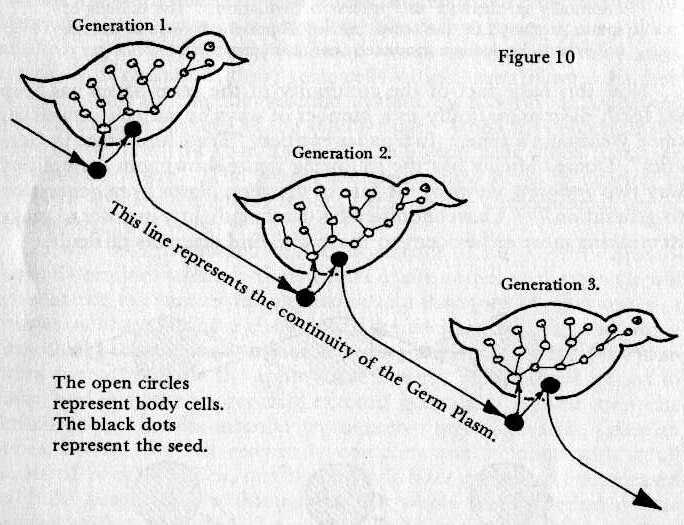
A more truly representative diagram
is that which Sir Alister Hardy presents in his This Living
Stream, re-drawn as Fig.10. (210) In this view the initial germ seed gives rise both
to the seed of the next generation and to the body. The seed
is shown after migration into the body that is to house it.
210. Hardy, Sir Alister, This Living Stream,
London, Collins, 1965, p.76.
pg.11
of 29
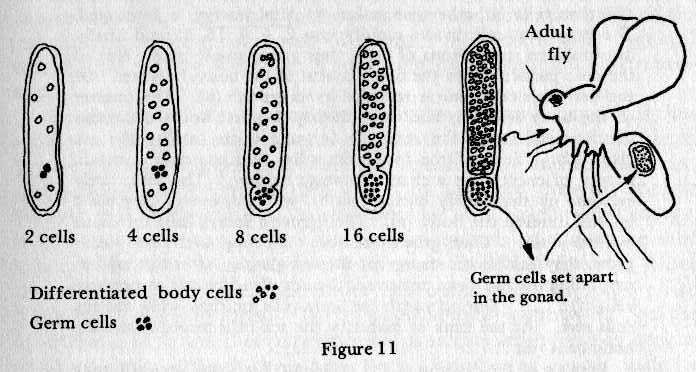
The next figure is re-drawn from an article written
by Fischberg and Blackler based strictly on experimental observation.
(211) The initial
germ plasm multiplies to the 8 cell stage by which time one or
two cells have been partially isolated and are shown as solid
dots. These cells, in the next stage, double and become effectively
isolated. In due time these isolated cells become the gonad germ
cells in the mature organism (in the Gall midge).
It may help, finally,
to bring this series of figures "home," as it were,
by re-drawing an illustration from Fritz Kahn in his Man in
Structure and Function, in which the same basic pattern of
continuity is transferred to the human context. (212)

Fritz Kahn published
his book about 35 years ago and it might be thought that it would
now be seriously out of date. In his discussion of this diagram,
however, the facts remain essentially as he has described them.
His work is still a very useful textbook, and the illustrations
are both imaginative and effective for the communication of what
is highly complex. He explains his diagram as follows:
211. Fischberg, M. and A. W. Blackler, "How
Cells Specialize," Scientific American, Sept., 1961,
p.134.
212. Kahn, Fritz, Man in Structure and Function, New York,
Knopf, 1960, vol.2, p.704, 705.
pg.12
of 29
The life
of man begins at the moment when the paternal sperm cell unites
with the maternal egg to form the egg-sperm cell that is the
conceptus (a). Through the union of the two chemically different
cells an otherwise unknown vital energy is developed. The egg-sperm
cell divides rapidly into 2, 4, 8, 16, 32 and finally into millions
and billions of cells, thus giving rise to man. Not all the cells
participate in the development of the body, however. Of the first
four cells, one is retarded in its growth (b). This quarter of
the body which is laid aside during the first hours of human
development forms the sex cells (c), and in its totality the
sex gland (d). Each of the four primordial cells possesses a
certain supply of energy like a charged storage battery. The
three body cells use up this energy during youth: with this energy
they build up and develop the body (e). Their growth energy lasts
for about twenty years. Then growth ceases. As long as the body
cells grow, they inhibit the energy of the sex gland. After this
inhibitory influence has been removed, the sex gland begins to
produce cells (f). . . . During youth the body cells multiply
while the sex cells rest. At the time of maturity the sex cells
multiply and the body cells rest. . . .
Because of the division of the
body into body and sex cells, man is not a unitary organism,
but rather a kind of double creature, consisting of himself and
his sex cells. . . . The sex cells do not belong to us
(as individuals) but rather to the entire species. . . . The
sex cells are the truly immortal element in us, as Plato already
sensed when he wrote, "The mortal creature harbors an immortal
element. . . ."
It may be difficult
to believe that any line of mortal creatures could convey
from generation to generation a continuing stream of immortal
cells without ultimately corrupting them. But evidently this
really is what takes place. C. E. McClung tells us: (213)
A germ cell of one individual
generation . . . becomes detached and forms a complete organism
of the next generation. By some insulating device [emphasis
ACC] the germinal elements within the gonad do not participate
in the somatic processes, but merely perpetuate themselves. [But]
on being freed from this inhibition [the insulating device] they
are freed from the limited role of mere germ cells and may perform
through their descendants all somatic activities.
But by nature,
and if not experimentally interfered with, these germ cells are,
as V. H. Mottram put it, "the only physically immortal things"
in our bodies. (214)
Again, I would modify this statement
slightly by noting that it is really the sex cells in the
female line that are "the only physically immortal things."
The statement does, however, show how widely it is recognized
that at the very root of our individual existence there is an
immortal constituent, the seed of the woman.
Weismann, almost a century ago,
in an essay published originally
213. McClung, C. E., quoted by Susanne Langer,
Mind: An Essay on Human Feeling, Baltimore, Johns Hopkins
Press, 1967, vol.1, p.408, footnote.
214. Mottram, V. H., The Physical Basis of Personality, London,
Penguin, 1949, p.25.
pg.13
of 29
under the title Uber
die Ewigheit des Lebens, i.e., "Upon the Eternal Duration
of Life," wrote (as we have already notedin chapter 1),
"The immortality of the unicellular organism has passed
over only to the ova or spermatozoa, the other [cells] must die,
and since the body of the individual is chiefly composed of them,
it must die also." Again, I would only modify Weismann's
statement by excluding the spermatozoa.
So the seed of the woman dies with
the woman because it is thus robbed of its home, even as seeds
die month by month if not fertilized by the sperm, being rejected
from the female body and to all intents and purposes killed in
the process. In any case they do not die because it is their
nature to do so or because they have limited energy. It is, as
we have seen, remarkably easy in animals to trigger the seed
into mitotic activity and thus to perpetuate itself indefinitely.
Once so stimulated, the ovum will under appropriate conditions
go on to full term in a viable form. There is considerable controversy,
on the other hand, as to whether parthenogenesis in this sense
has ever occurred in a woman, though there have been a number
of claims made by unwed mothers with respect to the birth of
supposedly fatherless daughters. (215)
Now all this has a direct bearing
upon the present theme. Having been endowed with immortality,
Adam and Eve acquired mortality. And, which is significant
from a physiological point of view, their offspring inherited
this acquired character. We have here therefore a clear case
of something which actually happened that, until comparatively
recent times, was ruled out as an impossibility. It has been
the traditional wisdom among geneticists for about a hundred
years that acquired characters are not inherited. Yet here was
an acquired character that "passed upon all men" (Romans
5:12). It seems rather strange to me that no Christian biologist
has given much thought to the matter. Indeed, it has taken a
man actually opposed to the Christian view to note this unusual
circumstance. Sir Gavin de Beer, an outstanding evolutionist
in England, when reviewing the book Mankind Evolving by
Theodosius Dobzhansky, makes this remark: (216)
One wonders if Pauline theologians
realize that the doctrine of original sin involves the inheritance
of an acquired character, for only genes can be inherited and,
by the nature of the case, neither Adam nor Eve when they first
appeared on the scene possessed the character they are alleged
to have transmitted to all their descendants.
To all their
descendants, save ONE! And here, if Sir Gavin had taken
the thought seriously, is a further great truth which might have
provoked him to think even more deeply upon the subject. For
it is
215. See Notes at the end of this chapter
(page 26).
216. de Beer, Sir Gavin, reviewing Theodosius Dobzhansky, Mankind
Evolving in Scientific American, Sept., 1962, p.268.
pg.14
of 29
evident that the acquired
character of Eden was indeed transmitted and must therefore
have ultimately reached the germ plasm. The only mechanism which
will satisfy all the conditions thus laid down in Scripture is
one which assumes that the transmission was effected via the
male seed only. Luther and Calvin and now Barth have all
recognized this fact. De Beer's comment is therefore not entirely
correct.
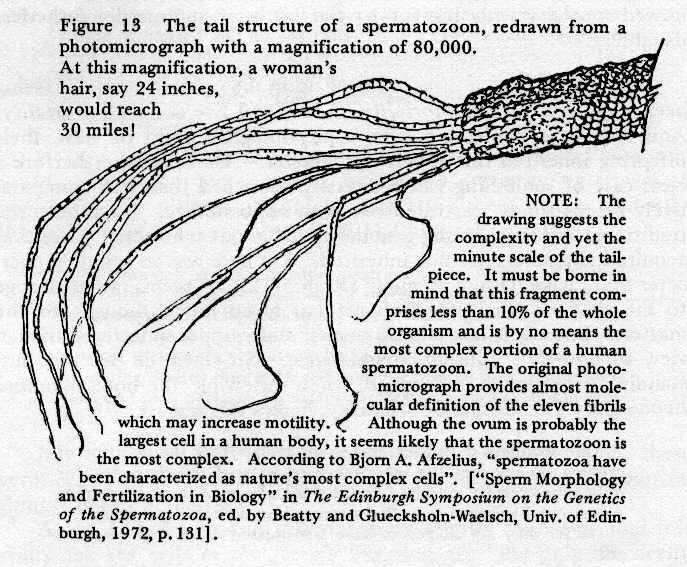
Now each spermatozoon is a single-celled
organism of highly complex structure and form. It is not only
the smallest cell in the body but quite possibly also the most
complicated in its organization. In its head-piece, it has a
nucleus containing the genes, suspended in a pool of surrounding
cytoplasm containing a number of minute structures of various
kinds called organelles (i.e., tiny organs), the whole
being sheathed in a membrane like a soft shell, and provided
with a quite complex tail portion by which a high degree of motility
is achieved. Fig.13 gives a good idea of its complexity.
It is now believed
that within the cytoplasm and among the minute particles which
are suspended in it, there are certain carriers of hereditary
pg.15
of 29
material which have
been termed plasmagenes. (217) These cytoplasmic "genes"
are distinct from the nuclear genes which hitherto have been
assumed the sole carriers of heredity, and they appear to be
(unlike the nuclear genes) susceptible to influences outside
the cell. Since there is constant interaction between the nucleus
and the cytoplasm of each cell it is possible for environmental
influences, by this route, to reach the hereditary material and
effect modifications in all subsequent generations. It is possible
that the fact of the continuous production of mature spermatozoa
throughout the adult fertile life of the male may expose them
to such influences in a way that the ova are not.
In the male the primordial germ cells
are present from the very beginning (as in the female embryo)
but the complex free-living organism which is the mature spermatozoon
(short-lived though it is) is continuously being manufactured
from puberty and throughout adult life by extensive modification
of the germ plasm cells. (218) And these mature spermatozoa are produced in the
hundreds of millions. This fact may account for the greater accessibility
of these cells to influences from the body which manufactures
and houses them. And the fact that the spermatozoon actually
penetrates into and becomes absorbed in and fused with the ovum
makes it a potential pathway into the ova for male body cell
influences.
Thus although the woman may have been
the first to introduce the fatal poison into her body cells,
she did not by that act poison her own seed, but the poison of
death does enter through the male seed into the seed of the woman
by the fusion of the two. By such a mechanism the poison in Adam's
body may have reached his seed, and via the cytoplasm of the
seed the poison is by fusion with the female seed passed on to
the embryo.
Now the substance of what we have
been discussing in this chapter may be stated as follows. As
we trace the history of biological theory with respect to the
mechanism of inheritance, we find Lamarck arguing that any animal
which responded to the challenges of the environment by developing
structures or instincts or chemical responses which gain for
it an advantage in the struggle to survive, was in a position
to further the chances of survival of its descendants by passing
these gains on by inheritance. The key doctrine here was that
acquired characters were inherited. It seemed self-evident and
necessary that this should be so in view of the apparent progress
of life, and it very reasonably accounted for the steady improvement
in the breeding of animals of particular interest to man. It
only remained therefore to unravel the mechanism whereby acquired
characters were transmitted.
217. See Notes at the end of this chapter
(page 27)
218. Hancock, J. L., "The Sperm Cell," Science Journal
June, 1970, p.32.
pg.16
of 29
It soon became apparent that this obvious "fact"
was not true after all. Acquired characters did not seem to be
inherited, or if they were, the mechanism was certainly
not a simple one. Once this was acknowledged, all kinds of every
day illustrations sprang to mind and made the older Lamarckian
view seem patently absurd: mutilated parents do not bear mutilated
offspring � daughters of Chinese mothers whose feet had been
bound from childhood bore normal daughters, circumcised fathers
did not beget circumcised sons, the blacksmith could have as
many puny infants as anyone else. A new law was therefore announced:
"Acquired characters are not inherited." And
all biologists accepted this new law at its face value.
A few biologists with Christian
convictions were disturbed by the new "law" because
they could see that real problems were created in our understanding
of the events which occurred in Eden. Mortality was acquired
by man, yet it was inherited. To quote Romans 5:12
again, "Death entered . . . and passed upon all men."
This was an essential aspect of the Fall of man and his need
for redemption. Was the Bible in error?
By the prodigious labours and elegant
methods of research of a number of geneticists and microbiologists,
the mechanism is now becoming clear. This research begins to
show that there are certain conditions under which an acquired
character can, after all, be inherited � not via the nuclear
genes but by something analogous to them in the surrounding cytoplasm
termed plasmagenes. The resistance to change in the germ plasm
is due to the fact that it is not derived from the body cells
� cells which are responsive to changes during life. It was
this fact that made it so difficult to see how the germ cells
could be influenced by what happens to the parents. It is the
plasmagenes that respond to influences, not the germ plasm.
But it now appears that although the
male germ cells, like the ova, are derived from the germ cells
of the parent body and not from the body cells, these male germ
cells are susceptible to the subsequent influence of the body
cells in a way that the female germ cells are not. The end result
is that by this roundabout way some acquired characters, whether
hurtful or harmless, seem to be inheritable in mammals through
the male seed. The pathway is from body cells to male germ cell
cytoplasm, and from the male germ cell cytoplasm to the
female seed by fusion at the time of fertilization. And thence
these modifications appear in body cells of the resulting offspring
both male and female. And these steps are repeated generation
after generation so long as the seed of the woman is fertilized
by the seed of the man.
But if the seed of the woman could
be activated without fertilization by the seed of man, it must
be supposed that the result would be the emergence of an individual
escaping the mortogenic factor
pg.17
of 29
which Adam bequeathed
via his seed to all subsequent generations. Such an offspring
would recover in his person the original physical immortality
with which Adam was endowed at his creation.
In short, to
summarize a long and complex chapter, it may be said that the
seed of the woman is the only remnant that has retained the original
immortality possessed by our first parents. By contrast, the
seed of man and the body cells of both the man and the woman
have been mortalized. Furthermore, even the seed of the woman
is fatally poisoned by fusion with the male seed.
However, this poison affects only that portion
of the woman's seed which will develop into body cells: the remainder
of her seed continues to form the immortal stream of germ plasm.
Only if an ovum from this germ plasm reservoir can be fertilized
by some means not natural to man can a body with the original
endowment of potential immortality be recovered again.
 pg.18
of 29
pg.18
of 29
NOTES
187. (See page 2) Many of the examples of acquired characters
that seem to have become inheritable which are now being discussed
by such men as Waddington, seem to me of dubious value because
they could be viewed equally well as preadaptations. This is
true of the thickening of the soles of the feet in the human
fetus. It was noted by Darwin and elaborated upon subsequently
by R. Semon [Arch. mikr. Anat., vol.82,1913, p.164ff.],
and it has since been discussed by C. H. Waddington in an article
entitled "The Evolution of Adaptations" [Endeavour,
12 July, 1953, p.136]. Among evolutionists, it is customary to
point to this phenomenon as having resulted from the bipedal
locomotion of man which has had the effect of toughening the
soles of his feet, an advantageous acquired character which is
then inherited after millennia of use. The human fetus now therefore
is born already prepared for walking, in this respect, according
to this view.
Waddington refers to a similar
situation in connection with the ostrich. This bird has two conveniently
located callosities on its breast which bear the brunt of friction
and pressure when the bird squats on the ground. According to
Waddington, these callosities have become inherited and they
are therefore found to be already formed during foetal development
["Experiments in Acquired Characteristics," Scientific
American, Dec., 1953, p.92f.]. However, this particular case
is not as straightforward as Waddington makes it appear, for
as Sir Gavin de Beer has pointed out, the ostrich is born with
other similar callosities which it cannot make use of at all
[Embryos and Ancestors, Oxford University Press, 1951,
p.87]. It could therefore be argued that we have here a case
of the accidental development of callosities due possibly to
some gene mutation, two only of which callosities happen to be
of some use to the animal.
A somewhat analogous situation
has been observed in man in the form of so-called squatting facets
of the Indians of Punjab. These Indians easily assume a restful
squatting position which the European finds difficult, because
of a modification of the bone structure of the tibia. No such
modification is ever found among chair-users, according to Wood
Jones [quoted by Kenneth Walker, Meaning and Purpose,
London, Penguin, 1951, p.154], but the Punjabis are born with
them. Is this, then, an acquired character that has become inheritable
or is it merely that they have made use of a chance modification
once they discovered its advantages?
Such proposed examples of inherited
acquired characters have, it seems to me, doubtful validity.
On the other hand, there is much experimental evidence on the
genuine inheritance of acquired characters in many forms of life
from the simplest to the more complex which seem most easily
to be accounted for by assuming that they are inherited cytoplasmicly
rather than via the nuclear genes. Some further observations
on this point will be found in a later reference, #217.
One of the most eloquent supporters
in recent times of what may be called Neo-Lamarckism was the
English naturalist, Professor F. Wood Jones. In his Trends
of Life, he has a whole chapter titled, "The Inheritance
of Adaptations," which is well worth examining [London,
Arnold, 1953]. And in the same year, Dr. Carlos Monge reported
an impressive example of what seems clearly to be a case of an
acquired character being inherited in man. Monge found that Andean
highlanders had developed considerably larger chests, presumably
a compensation for the rarefied atmosphere in which they live.
The interesting thing is that many of their descendants who came
down and have now lived along the sea-coast for many generations,
still have the same large deep chests and broad shoulders of
the highlanders. If this were simply a superficial response of
the highlanders to the need for an increased lung capacity, one
would expect it to disappear quickly in their lowland descendants.
That it has not done so, seems to indicate that the character
became inheritable ["Biological Basis of Human Behaviour"
in Anthropology Today, edited by A. L. Kroeber, Chicago
University Press, 1953, p.127ff.]. Why this lung enlargement
should become heritable but not the blacksmith's muscular build,
is hard to say. The mechanism is obviously not a simple one.
pg.19
of 29
191. (See page 3) Briggs, Robert and
Thomas King, "Nucleoplasmic Interactions in Eggs and Embryos"
in The Cell: Biochemistry, Physiology, and Morphology,
edited by J. Brachet and A. E. Mirsky, New York, Academic Press,
vol.1, 1959, p.539.
There is some evidence that some
of the body cells retain the full potential of the germ cells.
Writing in Science under the heading "Some Characteristics
of a Continuously Propagating Cell Derived from Monkey Heart
Tissue," J. E. Salk and Elsie N. Wood report that it has
been possible by the right techniques to isolate heart tissue
cells and induce them to go on multiplying indefinitely [Science,
vol.126, 1967, p.1338]. The phenomenon suggests that some of
the potential for immortality which is characteristic of germ
cells may have been retained even by the body cells which have
differentiated some distance from the originating germ plasm.
Recently MD of Canada reported
that Dr. John Gurdon and his co-workers at Oxford had grown fully
mature and fertile frogs from single body cells extracted from
the intestinal lining of other frogs. With his present technique
more than 30% of the intestinal cells could be made to grow at
least to the tadpole stage [vol.10, no.3, 1969, p.53]. Neither
lines of proliferating cells were human. It must surely be assumed
that the fall of man has made his body cells unlike all other
animal cells.
192. (See page 4) In a manner of speaking, Weismann was both
right and wrong in assuming that differentiating cells lose the
totipotency of the initiating ovum to the extent that such cells
are no longer individually capable of giving rise to a whole
animal but only to specific organs and tissues. In plants, of
course, the cells in a slip taken from almost any part of the
plant are capable of reproducing the whole organism, roots and
all. But experiments have now shown that complex animal forms
may also be reproduced by highly refined techniques from cells
which have long since differentiated into specific tissues and
have lost their identity as germ plasm.
The technique involves extracting
the nuclei from tissue cells and transferring them to enucleated
cells of germ plasm origin. Such reconstructed cells are evidently
capable of initiating the process of cell cleavage and division
and proceeding normally through embryological and foetal development
to maturity. It no longer seems likely, therefore, that cell
differentiation is due to the loss of gene material in the nucleus
during earlier stages of cell division but rather to changes
in the cytoplasm; although Briggs and King were able to demonstrate
that nuclei of cells taken from tissue which has formed later
in foetal development less frequently retain their totipotency
than do nuclei of cells derived from tissue formed earlier in
the developing embryo. It therefore seems likely that even the
nucleus may change slightly with time, although it is fairly
certain now that the major change occurring within the cell relates
to the chemistry or organization or structure of the cytoplasm
as successive cell divisions occur. The differentiated cytoplasm
interacts with the nucleus and this in turn leads to the emergence
of new directions for cell development along specific lines towards
the growth of tissues and organs which form the body or housing
for the original germ plasm.
Professor Bernard D. Davis, Harvard
Medical School, stated: "We now know that all the differentiated
somatic cells of an animal (those of muscle, skin, and the like)
contain in their nuclei the same complete set of genes. Every
somatic cell contains all the genetic information required for
copying the whole organism. In different cells, different sub-sets
of genes are acting while the remainder are inactive. Accordingly,
if it should become possible to reverse the regulatory mechanism
responsible for this differentiation, any cell could be used
to start the embryo. Though differentiation is completely reversible
in the cells of plants (as in the transfer of cuttings), it is
ordinarily quite irreversible in the cells of the higher animals.
The stability, however, depends on the interaction of the nucleus
with the surrounding cytoplasm. . . ." ["Prospects
for Genetic Intervention in Man," Science, vol.170,
1970, p.1280,1281].
pg.20
of 29
Cell
differentiation is therefore mainly the result of modifications
of the cytoplasm rather than the nucleus. A. C. Enders and S.
J. Schlafke, in a Ciba Foundation Symposium, observe that
the cytoplasm of cells, even by the time the blastocyst has formed,
is clearly different from the cytoplasm of the ovum. "During
the late cleavage stages and the blastocyst stage, the structure
of the cytoplasm alters a great deal in most species. Characteristically,
there is a diminution and re-organization of the cytoplasmic
inclusions. . ." ["The Fine Structure of the Blastocyst:
Some Comparative Studies" in Pre-implantation Stages
of Pregnancy, edited by G. E. W. Wolstenholme and M. O'Connor,
London, Churchill, 1965, p.45, 47]. Alfred Kuhn puts the matter
this way: "It is certain that the nuclei of some tissues
need not forfeit some of their talents to reach a certain stage:
rather they can replace the egg nucleus, and their derivatives
can satisfy all the demands of the developmental steps which
the various cells must pass through" [Lectures in Developmental
Physiology, translated by Roger Milkman, New York, Springer-Verlag,
1971, p.488]. It seems, therefore, that the cell nuclei retain
their totipotency to a far greater extent than the cytoplasm.
In the natural order of things, cells do fairly quickly become
differentiated and lose their totipotency � except perhaps
in plants. While most of the cells to which the totipotent ovum
gives rise soon become differentiated cytoplasmicly for the development
of body cells, not all of them do. A few remain for the perpetuation
of the germ cell line. It is these few that form the thread of
continuity from generation to generation.
The following readily accessible
articles dealing with this subject are useful: J. B. Gurdon,
"Transplanted Nuclei and Cell Differentiation," Scientific
American, Dec., 1968, pp.24-35; C. H. Waddington, "How
Do Cells Differentiate?", Scientific American, Sept.,
1953, pp.108�114; Michail Fischberg and A. W. Blackler, "How
Cells Specialize," Scientific American, Sept.,1961, pp.124�140;
Robert Briggs and Thomas J. King, "Changes in the Nuclei
of Differentiating Endoderm Cells as Revealed by Nuclear Transplantation,"
Journal of Morphology, vol.100, no.2, 1957, pp.269�311;
Lewis Wolpert, "Developing Cells Know Their Place,"
New Scientist, 14 May, 1970, p.322f.
195. (See page 6) Parthenogenesis is so well established for
so many species that it scarcely needs the reinforcement of this
note. However, for those who may not be aware of how widely it
has been demonstrated below man, the following brief comment
may be useful. Few proven cases of mammalian parthenogenesis
in nature have ever been clearly established, though as we have
already seen (ref. #175: see end Notes of Chapter 17) it has
been observed for lizards and is common enough among insects
and some fish. To the list of insects in which parthenogenesis
occurs naturally, B. I. Balinsky adds aphids, phyllopods, and
rotifers at certain times of the year, and of course bees in
which the fertilized egg produces a female and the unfertilized
egg develops into a male [An Introduction to Embryology,
Toronto, Saunders, 1970, 3rd edition, p.126].
The situation is very different
in the laboratory where experiment has shown that a very wide
range of animal forms can be induced to propagate parthenogenetically.
According to Albert Tyler, "Extensive investigations have
shown that in practically all the main groups of animals, normal
development can be obtained by artificial activation of eggs"
["Artificial Parthenogenesis," Biology Reviews,
Cambridge University, vol.16, 1941, p.292f.]. Reports include
such species as silkworms, caterpillars, sea urchins, star fish,
frogs, fish
pg.21
of 29
(including carp), lizards, birds and
rabbits. In 1896 R. Hertwig found that sea urchin eggs could
be activated by chloroform or strychnine! ["Ueber die Entwicklung
des ubefruchteten Seeigeleies," Festschr. fur gegenbauer,
Leipzig, 1896]. H. Spurway reports experimental parthenogenesis
in the guppy, Lebistes reticulatus ["Spontaneous
Parthenogenesis in a Fish," Nature, vol.171, 1953,
p.437]. In the case of rabbit ova cultivated in vitro, Dr. Chambly
in France almost fifty years ago was probably the first to demonstrate
that mammals can give birth to viable offspring parthenogenetically
[see Gregory Pincus, "Fertilization in Mammals,"
Scientific American, Mar.,1951, p.47]. There is some evidence
of man-induced parthenogenesis in sheep, though I am not sure
how dependable this is [Arthur Koestler, Beyond Reductionism,
London, Hutchinson, 1969, p.199].
196. (See page 6) "A human egg is a spherical cell .
. . which is one of the largest cells in the body, and when placed
against a dark background it is just visible to the naked eye.
. . . The large size of the egg cell is due mainly to deposits
of yolk in the cytoplasm. . . . In contrast to the egg, the sperm
is the smallest cell in the body. . . . The volume of an
egg cell is about 85,000 times that of a sperm" [Ursula
Mittwoch, Genetics of Sex Differentiation, New York, Academic
Press, 1973, p.84, 85].
197. (See page 6) It is established that an ovum can be activated
without fertilization by the spermatozoon, and in certain cases
will go on to full development of a mature female animal, complete
with a functioning reproductive system which thus provides the
initiating ovum with a mechanism for continuing itself indefinitely.
To this extent, the ovum is self-sufficient. The sperm does not
appear to be so, under natural conditions.
It was at one time supposed that
the limitations imposed upon the spermatozoon was entirely due
to lack of energy because of the small amount of cytoplasm surrounding
the nucleus. It simply starved before reaching sufficient maturity
to extract food from its environment. By reducing its food requirements,
at very low temperatures for example, its life can be greatly
extended, and certainly in warm-blooded animals the temperature
of sperm is quite critical to its survival, and unless the testes
descend to the scrotum free from the deep body temperature, they
are not viable. Excessive use of hot baths in Japan reduced male
fertility.
However, there appears to be some
other factor limiting sperm life. George Conner observed. "If
an ovum is cut into two pieces, one of which has no nucleus,
and the latter is then entered by a sperm, it too will divide
and become an embryo, though admittedly not as often as in the
case of the unfertilized ovum" [The Hormones in Human
Reproduction, New York, Atheneum, 1963, p.19]. This kind
of highly sophisticated manipulation of cells in the laboratory
is very different from anything that occurs in Nature whereas
the variety of treatments that can lead to parthenogenesis of
the ovum is so diverse that at least some of them must probably
occur under natural conditions.
Dorthea Rudnick, in her article
on Embryology in the1960 edition of the McGraw-Hill Encyclopedia
of Science and Technology [vol. IV, p.573], after pointing
out that the sperm itself is by no means essential for the activation
of the egg, suggests that the mature egg must be thought of as
a system containing all the potential factors for development
and the sperm essentially as a trigger that sets off the mechanism.
It is, of course, also the source of the paternal set of chromosomes.
But it seems clear that it is not at all the same
pg.22
of 29
kind of self-contained unicellular organism
capable of independent existence that the ovum is. Under natural
conditions, by itself it will die, whereas the ovum, left alone,
is in no necessity of doing so provided only that it is given
a suitable environment and an appropriate though remarkably non-specific
stimulation to activate it. If the ovum can thus survive by itself,
it can hardly be argued that the sperm contains any absolutely
essential component for its activation, since apparently the
ovum can be activated without it.
198. (See page 6) The ovum can be activated and developed
into a fully mature organism by an amazingly diverse range of
stimuli. According to Albert Tyler, "It is clear that there
is very little specificity in regard to activating agents. Thus
eggs of the sea urchin can be activated by such diverse agents
as puncture, heat, cold, ultra-violet radiation, radium emanation,
acids, bases, isotonic salt solutions, hypertonic and hypotonic
solutions, fat solvents and some alkaloids. This contrasts with
the high degree of specificity in fertilization (in nature)"
["Artificial Parthenogenesis," Biology Reviews,
Cambridge Univiversity, vol.16, 1941, p.318].
To these non-specific stimuli have
since been added others, including electric shock. The carp eggs
mentioned above (ref.#195, page 21) were actually activated by
human saliva. Professor Christian P. Raven of the University
of Utrecht has added to this growing list of activators such
physical treatment as illumination, induction shock and osmotic
pressure as well as chemical agents such as urea and saponin
[An Outline of Developmental Physiology, translated by
L. de Ruiter, New York, McGraw Hill, 1954, p.20].
I do not know whether I am reading
too much into the evidence, but I think it worth noting that
Balinsky observed that most of the agents used are of such a
nature that they probably damage the ovum to a greater or lesser
degree, and if applied too vigorously they cause the death of
the cell. He then adds, "It is reasonable to suppose, then,
that activation of the egg involves some kind of sub-lethal damage
to the egg cytoplasm" [An Introduction to Embryology,
Toronto, Saunders, 1970, p.127]. Possibly the human sperm fertilizes
(and so activates) the ovum but at the same time introduces some
type of damage to the cytoplasm which is not merely sub-lethal
but lethal. One need only hypothesize that this lethal effect
becomes operative only after the original single cell (the ovum)
has divided several times into a number of proportionally smaller
cells, the proportion of cytoplasm to nucleus accordingly being
reduced and presumably modified. It is known that with successive
divisions there is a progressive change in the amount of cytoplasm
as well as in its internal organization and its chemistry. The
nucleus meanwhile retains its original size and constitution.
Raven remarks: "The local concentration of certain substances
will initiate chemical reactions which previously were unable,
or almost unable, to take place because of the dilution of the
reagents or because of the presence of inhibiting substances"
[An Outline of Developmental Physiology, translated by
L. de Ruiter, New York, McGraw Hill, 1954, p.63]. Transferred
to the present context, this observation could very nicely point
the way to the actual mechanism whereby some contribution from
the cytoplasm of the sperm cell finally becomes lethal in its
effect on the growing organism.
A perceptive reader may discern
the importance of the finding of the non-specificity of the activating
agent. I think the introduction of the word activate is
significant in the present context because it would be such an
appropriate word to apply to the "overshadowing" by
the Holy Spirit (Luke 1:35). The preparation of the perfect body
that was to be animated for the Lord Himself was truly a miracle
in that a male child was born. But the stage was clearly
already designed for just such a tremendous event, when the event
is viewed in its physiological context. Nevertheless, it is important
to bear in mind that some creative power must still have been
at work to supply the Y chromosome in order that a man child
might be born, not a female child as would otherwise have been
the case.
pg.23
of 29
199. (See page 6) Where birds are concerned,
parthenogenesis is always found to result in males. This is because,
for some unknown reason, the sex determining roles of the X and
Y chromosomes have been reversed. Thus M. W. Olsen and S. J.
Marsden, in reporting on "Natural Parthenogenesis in Turkey
Eggs," note that in spite of the fact that there had been
no male contribution, all the parthenogenetic embryos carried
the diploid chromosome number and all the eggs which reached
a sufficient stage of maturity to allow for sex determination
were found to be male. Seventy-nine turkey hens were involved
in this experiment. Males were rigidly excluded. During the eight
week period in question, 2537 eggs were laid, of which 568 showed
parthenogenetic development. Forty-nine of these differentiated
to the extent that blood vessels were clearly visible. In 27
embryos were identifiable on gross examination. Four of these
allowed sex to be determined. All were male [Science,
vol.120, 1954, p.545].
Interestingly, Origen (c.185�c.254)
in his Contra Celsius [1,37] mentions that in his time,
vultures were reputed to raise parthenogenetic young.
201. (See page 7) It is often said that Weismann's views have
since been proven in error in certain important respects and
that it is therefore unwise to quote him in the light of what
we now know. This is unfortunate because the lucidity and insight
with which he presented his ideas makes them a most suitable
vehicle for communicating some very complex aspects of early
embryological events and his contribution to our basic understanding
has been tremendous in terms of the stimulus of his powers of
description. He appears to have been in error in one of his conclusions,
though the error has not otherwise damaged his general thesis
regarding the continuity of the germ plasm. In one other matter
he was mistaken but only in the sense that he visualized the
wrong mechanism for the right results. Thus his questionable
contributions are really limited to the two following points:
(1) the role of the second polar body or cell which is cast out
by the ovum in its earliest stages of preparation for the admission
of the sperm and (2) the mechanism by which body cells are differentiated
from germ cells, and the claim that only the germ plasm retains
all the developmental capability of forming a new organism. [See
his "Continuity of the Germ Plasm as the Foundation of a
Theory of Heredity" in Essays Upon Heredity and Kindred
Biological Problems, translated by E. B. Poulton, S. Schonland
and A. E. Shipley, Oxford University Press, 1889, vol.1, p.214,
225].
With respect to the role of this
second polar body, this small cell is expelled from the ovum
immediately prior to the penetration of the sperm, removing from
the ovum half its chromosomes and thus reducing it to a haploid
cell so that the contribution of the sperm nucleus will restore
it as a normal diploid cell rather than overburdening it with
supernumerary chromosomes. Weismann surmised that this polar
cell probably removed the male component of the heretofore hermaphroditic
ovum (no one knew anything at that time about the X and Y chromosomes)
and thus opened the way for the sperm's contribution to enter
unchallenged. It is not believed at present that this is so.
Weismann believed that the ovum
could conceivably be fertilized by this polar body rejoining
the ovum, or possibly even before it had broken away and gained
its independence. In any case, Weismann may not have committed
himself altogether to the view that the nucleus of the ovum was
hermaphroditic, but was merely stating that this was the opinion
of some of his contemporaries. He had in mind C. S. Minot and
F. N. Balfour in particular. The latter had said, "the function
of forming polar cells has been acquired by the ovum for the
express purpose of preventing parthenogenesis" [A Treatment
of Comparative Embryology, London, Macmillan, 1880, vol.1,
p.63].
pg.24
of 29
With respect
to his second point, we have already seen that the nuclei of
some somatic cells, contrary to Weismann's surmise, do apparently
contain the whole original complement of the germ cells, but
in most animal species the cytoplasm of these somatic cells does
not. There is therefore a real loss of potential in the cell
as a whole. Thus Weismann was effectually correct, only he was
wrong about the mechanism.
Bernard D. Davis notes in this
connection: "Every somatic cell contains all the genetic
information required for copying the whole organism. In different
cells different subsets of genes are active, while the remainder
are inactive. Accordingly, if it should become possible to reverse
the regulatory mechanism responsible for this differentiation,
any cell could be used to start an embryo" ["Prospects
for Genetic Intervention in Man," Science, vol.170,
1970, p.1280]. In some species body cells of some organs do seem
to retain the potential of acting as germ cells for a new organism.
In frogs, for example, it has been demonstrated that some body
cells have retained the totipotency of the germ cells. To this
extent therefore Weismann was mistaken in supposing that the
nuclei of the germ cells lost some of their character in the
formation of somatic cells. The nucleus of the germ cell loses
nothing except the power to express itself fully, surrendering
this totipotency in response to changes in cytoplasmic determinants.
This cytoplasmic change has been termed 'chemodifferentiation.'
As Robert Briggs and Thomas J. King conclude, the process of
organogenesis appears to be due to the localized development
"of certain cytoplasmic materials. . . . The zygote
nuclei play no specific role in this localization" ["Nucleoplasmic
Interactions in Eggs and Embryos" in The Cell: Biochemistry,
Physiology, and Morphology, edited by J. Brachet and A. E.
Mirsky, New York, Academic Press, vol.1, 1959, p.540]. Thus the
nucleus, whatever its potency per se, is evidently subservient
to the cytoplasm and will generate what the cytoplasm allows.
Balinsky points out that Weismann
has been chiefly criticized for his observation that it is the
"unequal fission of the nuclear material which introduces
differentiation in developing tissues" [An Introduction
to Embryology, Toronto, Saunders, 1970, p.139]. But one only
need correct this to read "unequal fission of the cytoplasmic
materials" instead of nuclear materials in order to make
it a perfectly valid statement. In the circumstances, this is
a small error and a remarkable tribute to Weismann's insight.
His basic concept of the continuity of the germ plasm remains
essentially unchallenged.
206. (See page 10) The number of cells which initially retain
the full potential of the germ plasm at the moment when the first
specialized cells begin to appear as body cells varies in different
species, but thus far it appears that it is remarkably small.
According to M. Fischberg and A. W. Blackler, the beginning of
differentiation of cells in the gall midge, Mayetiola destructor,
must be placed about the third division when there are only eight
cells ["How Cells Specialize," Scientific American,
Sept.,1961 p.134].
208. (See page 10) Money, John and A. A. Ehrdardt, Man
and Woman, Boy and Girl, Baltimore, Johns Hopkins University
Press, 1972, p.37. The once active controversy as to neogenesis
(the origination of oocytes de novo) in post-natal life
seems to have been laid to rest by a general consensus of opinion
that there is an unrenewable stock of oocytes at birth. On this
point, see P. L. Krohn, "The Biology of Aging," Nature,
11 Jan.,1958, p.74; and Gregory Pincus, "Reproduction"
in Annual Review of Physiology, vol.24, 1962, p.57. Ursula
Mittwoch observes, "Cell division in human oogonia occurs
only during foetal life. . . . At the time of birth the oogonia
have already been transformed into primary oocytes" [Genetics
of Sex Differentiation, New York, Academic Press, 1973, p.87].
pg.25
of 29
215. (See page 14) An editorial comment
in the world renowned medical journal, Lancet, under the
heading, "Parthenogenesis in Mammals?" has this observation:
"The possibility that a woman might become pregnant without
at least one spermatozoon having entered the uterus is not one
which 'reasonable men' would likely entertain. Scientific opinion
for several centuries has sided with the reasonable man; but
today, biologists and cytogeneticists in particular would be
less dogmatic in discussing such a possibility" [5 Nov.
1955, p.967].
The writer then notes the number
of species of animals in which parthenogenesis has been observed
to have occurred in nature or under laboratory conditions, and
asks, "In view of this, we may have to re-examine the justification
for our belief that spontaneous parthenogenesis is rare in vertebrates
and absent in mammals. If it were rare in mammals but occasionally
present would it in fact be noticed?"
Subsequently he observes further:
"There are sound reasons which might lead biologists, if
not to expect parthenogenesis occasionally in mammals, at least
to look out for it." He suggests that "man is clearly
the mammal in which parthenogenesis would be least likely to
pass unnoticed." But I submit that this is a questionable
assumption. I think it was Sir Peter Medawar, Director of the
Medical Research Council in England, who said that the claim
by an unwed mother of a fatherless child would not be believed,
and in a married woman would not be noticed. I think he was correct,
human nature being what it is. But the editorial comment does
point out that immunological make-up of a parthenogenetic child
would be such that "it could be recognized with absolute
certainty" by appropriate medical techniques. The point
is that being so born, such a daughter would theoretically share
the genetic constitution of the mother completely, and it should
be possible therefore to make a skin graft from the daughter
back to the mother with complete success. There would be other
possible tests of a like nature also.
Subsequent correspondence stirred
up enough interest in England at the time that an invitation
went out publicly to any woman who believed she had given birth
to a parthenogenetic daughter to submit to supervised medical
examination along such lines. Later correspondence in Lancet
of June 30 of the following year (1956) indicated that nineteen
corespondents had presented themselves for examination.
Dr. Helen Spurway, Lecturer in
Biometry and Eugenics at University College, London, was invited
to conduct these tests. And to make a long story short, of these
nineteen pairs of mothers and daughters, eleven were eliminated
after interview for various reasons, and blood tests eliminated
another four. Of the remaining four, three were eliminated due
to such factors as incompatible eye or hair colour, etc. This
left only one mother-daughter pair as a genuine test case.
A battery of tests were conducted
between these two which, for the most part favoured the mother's
claim. However, a skin graft from the daughter to the mother
began to show signs of rejection after six weeks and was later
easily wiped off with damp cotton wool. Spurway points out that
the daughter would not be completely genetically identical with
the mother, since during meiosis the ovum would have made
a selection of only 50% of the original genes in the mother's
body cells and the daughter could not therefore have a completely
identical constitution. It is possible, as a consequence, that
even this test may not be sufficiently valid by itself.
pg.26
of 29
Spurway
concluded, therefore, that "rigorous proof is impossible,
though it remains true that all the evidence obtained from serological
and special tests is consistent with what would be expected in
a case of parthenogenesis." As she noted, the absence of
any knowledge on the woman's part of such a possibility "adds
to the probability of such a claim being well founded. This mother's
claim must not only be considered seriously but it must also
be admitted that we have been unable to disprove it."
217. (See page 16) C. L. Prosser observed: "Several types
of non-genic inheritance and of indirect effects of environmental
selection on the genotype are recognized. Cytoplasmic inheritance
is being discovered in more and more groups of organisms, and
cytoplasm is more readily influenced by environment than is the
nucleus." ["The Origin After a Century: Prospects
for the Future," American Scientist, vol.47, 1959,
p.545]. He instances cytoplasmic particles which may be transmitted
� for example, the granules for the Kappa factor in paramecium
or the plastidids of certain plants; also cytoplasmic factors
are involved in the inheritance of serotypes in ciliates.
In his small but challenging and
lucidly written little book, Nucleo-Cytoplasmic Relations
in MicroOrganisms [Oxford University Press, 1953], Boris
Ephrussi underscored the fact that body cells can be made to
"breed true for a practically indefinite time." We
have seen this, of course, in connection with Alexis Carrel's
chicken heart tissue [see Chapter 1]. We have already noted that
Hayflick's apparently contradictory data may have been at fault
through cultivation in an inadequate medium [see ref.#123, Part
I, chapter 8]. Ephrussi therefore argues that since such cells
have precisely the same nuclei that the originating fertilized
ovum possessed, these lines of body cells ought to begin to revert
to the undifferentiated ovum type of cell if the nucleus alone
is providing the blueprint for their development. It has to be
concluded, therefore, that these body cells do not revert because
some other inheritable blueprint exists in the cell apart
from the information in the nucleus. Such information must
be contained in the cytoplasm. So muscle cells replicate as muscle
cells, for example, because the cytoplasm is instructing them
to do so and not because of guidance provided by the nucleus.
That muscle cells do not replicate suddenly as some other kind
of cell, forces us to believe that each kind of tissue is formed
of cells with a unique cytoplasm. But this cytoplasm can be acted
upon by the environment and changed in its constitution so that
a given type of cell may begin to build a new type of tissue.
This has a bearing, probably, on the phenomenon mentioned by
Sir Peter Medawar in which a completely functioning ball and
socket joint may form in an entirely exceptional location [The
Art of the Soluble, London, Methuen, 1967, p.26].
In view of the fidelity with which
particular tissue cells will go on replicating themselves, Ephrussi
says: "Here we have identical perpetuation, but in this
case the inherited differences can hardly be ascribed to nuclear
genes, for the different cell types which make up (the organism)
are all derived from the egg cell by equational mitosis. They
must therefore all possess the same genotype. . . . The
differential must have its seat in the cytoplasm" [op.
cit., p.4].
Ephrussi then proceeds to detail
the work done with unicellular paramecia which multiply by conjugation,
showing clearly that they can pass on certain modifications (in
particular, a kind of mortogenic or "killer" factor)
not via nuclear genes but via plasmagenes. As he says: "These
studies confirm the view that the cytoplasm, like the genes,
is endowed with genetic continuity. The genes are therefore no
longer to be regarded as the sole cell-constituent endowed with
this property" [p.6].
pg.27
of 29
Ephrussi
sums up his conclusions thus: "Considering that embryonic
development results in a restriction (and some widening, too)
in different cell lineages of the manifold potentialities originally
carried by the egg, we may picture the process of differentiation
as consisting, for example, in the segregation or sorting out,
of an initially mixed population of cytoplasmic particles. Or
we may suppose that the egg, to begin with, contains a mixed
population of inactive particles, and that development consists
in the activation by nuclear genes of different sorts of lineages"
[p.100].
If we assume the former, it is
clear that some foreign substance entering the germ cell cytoplasm
at some critical stage of its internal organization, could effect
a change that in due time will rob it of its power of endless
self replication � will, in short, rob it of its potential
immortality. This poison may have gained entry in the cytoplasm
of the fertilizing spermatozoon. But there is also another possible
route.
B. Baccetti and B. A. Afzelius
have recently published a work on the Biology of the Sperm
Cell [See ref. #122 in Part I, chapter 8] which reflects
the ever growing interest in the cytoplasm, by contrast with
the preoccupation with the nucleus which has tended to prevail
for some years. They speak of the mitochondria in the sperm cytoplasm
which appear to have proteinaceous crystal inclusions that may
account for as much as 50% or more of the sperm volume in some
species. There is as yet no manifest function for these crystals
except possibly as a source of nutrition for the fertilized ovum,
somewhat analogous to the yolk granule.
Since the sperm mitochondrion is
small compared with the egg volume, and since normally only one
sperm, or at the most a few spermatozoa penetrate the egg, the
idea seems far-fetched. On the other hand, the spermatozoa that
are not taken up by the fertilized ovum perish by the hundred
million in the vicinity of it and their cumulative proteinaceous
crystal may in the aggregate form a substantial reservoir of
nutrient.
We thus have here a situation where
the quantity of sperm cytoplasm available to the growing ovum
may be quite significant in its influence on the cytoplasm of
the ovum, by absorption from its immediate environment. It may
well play a significant role in its early stages of development,
and it is these early stages of development � the first four
or five doublings of the zygote � that the germ line cells
are set aside and the body cells begin their differentiation.
There is increasing evidence of
the critical importance of the cytoplasm and of changes in its
composition after successive cleavages, as soon as the embryo
has reached the stage of development which witnesses the initiation
of cell differentiation and the formation of body cells as opposed
to the mere replication of germ plasm.
A. I. Caplan and C. P. Ordahl have
now proposed, as the result of a very elegant series of experiments,
that, "The expression of those genes necessary . . . to
give rise to diverse cell types is wholly dependent on exposure
of cell nuclei to a small portion of egg cytoplasm. The general
state of the cell and the activity of the cell's cytoplasm provide
important signals for the developmental programs. . . . The
segregation of cytoplasm during cleavage establishes extranuclear
environments that are determinants to the developing organism"
["Irreversible Gene Repression Model for Control of Development,"
Science, vol.120, 1978, p.120�130].
It is possible therefore that the
poison may remain quiescent and effectively neutralized in the
multiplying cells of the fertilized ovum until the cleavage has
proceeded to the point of either eliminating a certain quantity
of cytoplasm by reduction in cell size, or modifying certain
cytoplasmic factors that had hitherto served as neutralizers.
Thereafter, the progressive change in the composition of the
cytoplasm subjects the cell increasingly to the original protoplasmic
poison which renders all body cells mortal as a consequence.
pg.28
of 29
Baccetti
and Afzelius point out in their study that many human spermatozoa
have bizarre shapes and these are probably the results of faults
in the spermiogenesis. For each particular man a sperm sample
will show many differently shaped, abnormal spermatozoa, the
percentage of which is characteristic of that individual and
apparently constant throughout the years. Moreover, a number
of diseases such as the common cold can cause a temporary increase
in the number of abnormal sperm cells. This demonstrates the
fact that the sperm are susceptible to damage from their environment.
The authors believe that in spite of the high incidence of defective
or abnormal sperm, they actually have little consequence for
fertilization itself unless the proportion of normal sperm is
exceptionally low. For the defective sperm do not reach the ovum:
or they fail to penetrate it. "The many defective spermatozoa
are somehow prevented from attaining the fertilizing site"
[The Biology of the Sperm Cell, Monographs in Developmental
Biology, #10, Basel, Karger, 1976, p.19,158,160].
Thus within the developing ovum
in its initial stages, two lines of cells soon appear: germ cells
and body cells. The determinant that brings about subsequent
differentiation of body cells in accordance with their destiny
as various organs and their loss of potency as germ cells, appears
to reside in the cytoplasm. When the germ line cells are set
aside, it is by a kind of quarantine which permits the body cells
to be progressively differentiated by factors in their cytoplasm.
The process of differentiation evidently exposes them to some
mortogenic factor derived perhaps from their source of nourishment.
At any rate there seems little doubt that the differentiating
body cell lines are established and maintained by cytoplasmic
determinants, because the nuclei in all the cells (germ and body
alike) remain unchanged.
That unicellular forms (amoeba
for example) which are "by nature" immortal may be
mortalized by a change in diet has been demonstrated repeatedly.
And the mortalized amoeba are now altered in the composition
of their cytoplasm, not in the composition of the nucleus. If
mortalized amoeba cytoplasm is transferred to immortal amoeba,
the latter become mortalized ["How Immortal Are Amoeba?",
a note in Nature, vol.217, 1968, p.706, 707]. There therefore
seem to be now in operation all the mechanisms we require, theoretically,
to account for the situation that exists in the human species
for whom physical death comes as an unnatural termination
to life.
It might be thought that if animals
die and the process of fertilization is very similar, must we
not then suppose that the same poison in their case is having
the same effect via a similar route? Then do animals die for
the same reason that man does . . . because they, too, have somehow
been poisoned? The answer to this may, I think, lie in one of
two directions. First, we do not know for certain that death
is inevitable for animals, except as a result of "accident."
It is statistically certain that every animal will die but it
may not be inevitable biologically, as we have seen in Part I;
and it is, in my view, certainly not a penalty. So that in the
sense in which we speak of death as having "entered"
(Romans 5:12) for man as a result of the Fall, we have no necessary
reason to suppose that death as penalty has "entered"
into the stream of animal life. The second point is that
we do know that Adam and Eve were not subject to death at first.
What we may only surmise about animals, namely, that they have
a potential for physical immortality, we may know with considerable
assurance about man from what we are told in Genesis. He was
"killed" by ingesting a forbidden fruit. Thus for man
we might expect to find, and I believe we should seriously look
for, the mechanism whereby mortality as an acquired character
has been transmitted. There is no particular reason to assume
that such a mechanism exists among other living things: indeed,
we know that for millions of living things such a mechanism does
not exist, for they are truly potentially immortal.
pg.29
of 29
Copyright © 1988 Evelyn White. All rights
reserved
Previous Chapter Next
Chapter
|
